
Bed net and bed bug dimensions
The three bed nets we used differed in their insecticide treatment and the size of mesh holes through which bed bugs would need to pass. The untreated bed net (Siam Dutch) had relatively symmetrical holes measuring 1.39 ± 0.05 mm by 1.38 ± 0.13 mm (n = 20). The deltamethrin-treated bed net (PermaNet) was similar in hole dimensions to the untreated bed net (1.65 ± 0.04 mm by 1.33 ± 0.07 mm, n = 20). The permethrin-treated bed net (Olyset) had much larger oblong holes, measuring 3.78 ± 0.25 mm by 1.95 ± 0.24 mm (n = 20) (Fig. 3). Analysis of the nets revealed that hole lengths were significantly different across the three bed nets (F = 1333.07, df = 2, 57, P < 0.0001) (Fig. 3). Only the Olyset net had significantly wider holes than the other two nets (F = 102.91, df = 2, 57, P < 0.0001). Therefore, based on hole dimensions alone, we expected the bed nets with smaller holes to selectively allow only smaller bed bugs to pass through, whereas bed bugs should be least constrained passing through the Olyset net.
Fig. 3
Comparisons of the dimensions of mesh holes in bed nets used in this study. The width (orange) and length (blue) of 20 randomly selected holes were measured for each of the three bed nets. The mean is represented by x within each box plot. Photos of each net are shown with the same scale bars (1 mm). The widths of bed nets that share lower case letters are not significantly different from each other, and the lengths of bed nets that share upper case letters are not significantly different from each other (ANOVA, Tukey’s HSD, P > 0.05)
We used a total of nearly 3800 bed bugs in the assay validation and data collection phases of this project. We measured the maximal thoracic and abdominal widths of various developmental stages of unfed and fed bed bugs of both strains (Fig. 4). Overall, the maximal thoracic and abdominal widths of fed bed bugs were similar to the respective widths of unfed bed bugs. In some stages (e.g. second instars, adults), fed bed bugs were significantly narrower than unfed bed bugs, indicating that the enormous increase in body mass associated with full engorgement on a blood meal resulted in longitudinal stretching of the abdomen and an increase in its girth that stretched the pleural membrane, decreasing the width of the abdomen. The mean thorax width of Harlan strain bed bugs ranged from 0.49 ± 0.01 mm in fed second instars to 2.12 ± 0.03 mm in fed adult females (n = 20 for each stage) (Fig. 4a) and in Fuller Mill bed bugs from 0.80 ± 0.02 mm in unfed second instars to 1.97 ± 0.03 mm in fed adult females (n = 20 for each stage) (Fig. 4b). Mean abdominal widths in Harlan bed bugs ranged from 0.88 ± 0.03 mm in fed second instars to 3.24 ± 0.03 mm in unfed adult females and in Fuller Mill bed bugs from 1.06 ± 0.03 mm in unfed second instars to 2.86 ± 0.03 mm in unfed adult females.
Fig. 4
Comparisons of the thoracic and abdominal widths of unfed and fed C. lectularius of two strains. The thoracic (Tx) and abdominal (Ab) widths of second, third and fifth instars, as well as adult males and females (20 bed bugs per stage), were measured at both unfed (orange) and fed (blue) states for both the Harlan (insecticide-susceptible, a) and Fuller Mill (pyrethroid-resistant, b) strains. The results show increases in both thoracic and abdominal widths throughout development and general decreases in abdominal width in fed bed bugs. The mean is represented by x within each box plot. The average largest linear dimension of mesh holes in the three bed nets are represented by red dashed lines. Significant differences (Student’s t-test) in width between fed and unfed bed bugs are denoted by *P < 0.05; **P < 0.01; ***P < 0.001
In general, the Harlan and Fuller Mill bed bugs broadly overlapped in their thoracic and abdominal widths. The thoracic widths of fed second instars and adult male Harlan bed bugs were significantly smaller post-feeding (Student’s t-tests, t = 13.16, df = 38, P < 0.001; t = 2.39, df = 38, P = 0.0219). Harlan second instars, as well as adult males and females, had significantly smaller abdominal widths after feeding (Student’s t-tests, t = 3.10, df = 38, P = 0.0042; t = 5.46, df = 38, P < 0.001; t = 5.17, df = 38, P < 0.001). Interestingly, the width of the thorax in Fuller Mill bed bugs did not change significantly in any life stage, whereas abdominal widths were significantly smaller in fed than unfed adult males and adult females (Student’s t-tests, t = 5.11, df = 38, P < 0.001; t = 6.36, df = 38, P < 0.001); the reverse was evident in second instars, with larger abdominal widths in fed nymphs (Student’s t-test, t = 2.50, df = 38, P = 0.018).
To estimate which bed bug stages might be excluded by each of the three bed nets, we statistically compared the maximum widths of each stage of unfed and fed bed bugs, respectively, with the hole lengths of each bed net. Resulting significant differences in size were associated with either a predicted ability to pass through the respective net (smaller size) or a predicted exclusion by the net (larger size), independently of the effects of insecticides. For brevity, only those life stages around the “cut-off” point, where exclusion or passage may be based on feeding status, as determined by statistical comparison, are presented. In the case of the untreated (Siam Dutch) bed net, we found that only second instars of both Harlan and Fuller Mill were significantly smaller than the measured hole length (F = 596.51, df = 40, 779, P < 0.0001; Dunnett’s test, P < 0.05), and therefore both fed and unfed second instars were predicted to pass through this net. Interestingly, most third instars of both strains were predicted to pass through unfed (Dunnett’s test, P < 0.05), but might potentially be excluded once fully fed (Dunnett’s test, P > 0.05) (Fig. 4). Similar results were seen with the deltamethrin-treated (PermaNet) net, as second instars of both strains, as well as third-instar Harlan bed bugs, were expected to pass regardless of feeding status (F = 606.38, df = 40, 779, P < 0.0001; Dunnett’s tests for both, P < 0.05). However, most third-instar Fuller Mill bed bugs, as well as most fifth instars of both strains, were predicted to be excluded once fully fed (all three Dunnett’s tests, P > 0.05) (Fig. 4). In the case of the permethrin-treated (Olyset) net, all measured life stages of both strains were significantly smaller than the mesh hole length, so most were predicted to pass through the net regardless of feeding status (F = 674.45, df = 40, 779, P < 0.0001; Dunnett’s tests for all, P < 0.05) (Fig. 4).
Blood meal-seeking assays with multi-stage bed bug cohorts
To empirically investigate the predictions of statistical tests, we assayed mixed cohorts of Harlan strain bed bugs of all previously measured life stages to identify which would pass through the nets and which would not. We compared the untreated Siam Dutch bed net, with small mesh holes and no interference from insecticides, and the permethrin-treated Olyset bed net, with large mesh holes. In the blood meal-seeking assays, second, third and fifth instars readily passed through the untreated bed net, with mean feeding percentages of 100%, 98% and 94%, respectively, whereas 82.7%, 86% and 96% passed through the permethrin-treated bed net and blood fed (Fig. 5). Due to the large variation among replicates with the permethrin-treated bed net, we found no significant differences in the responses of second, third and fifth instars on the untreated and permethrin-treated bed nets (P > 0.05), which generally aligns with our statistical analysis of net hole length and bed bug size. Of note, however, is that while the passage of second and third-instar unfed bed bugs through the untreated net was expected based on our prior analysis, the passage of unfed fifth instars was not. This potentially relates to the pliable nature of the nets, which in certain cases might allow bed bugs to pass through successfully, as well as the more pliable nature of the bed bug abdomen than the thorax.
Fig. 5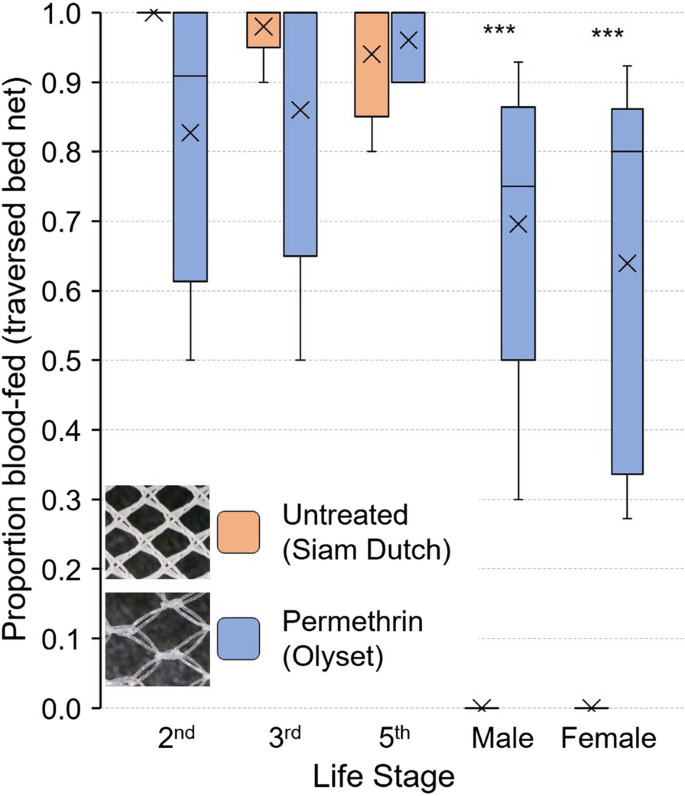
Comparison of the blood meal-seeking assays of insecticide-susceptible Harlan strain C. lectularius bed bugs of various life stages through the Siam Dutch untreated bed net and the Olyset permethrin-treated bed net. The mean is represented by x within each box plot. No significant differences were seen between the two bed nets in the proportion fed (traversed the bed net) for second, third or fifth instars. However, significantly more males and females passed through the permethrin-treated bed net (larger mesh holes) than through the untreated bed net (Student’s t-test, ***P < 0.0001)
However, the untreated net completely excluded adult males and females from reaching the blood source (0% fed), whereas on average 69.6% and 63.9% of the males and females, respectively, traversed the permethrin-treated bed net and fed (Fig. 5). The two-way ANOVA analysis showed a significant effect of the overall model on bed bug blood feeding. Bed bug life stage, LLIN used and the interaction between stage and net significantly influenced the proportion of successfully blood-fed insects (Table 2). Specifically, the passage and feeding of adult bed bugs through the two nets were significantly different (Fig. 5) and aligned with the predicted results from our prior analysis. These results suggest that while bed nets with tight mesh (small holes) may stretch, and bed bugs may alter the width of their abdomen, these adjustments are constrained, as all adults were prevented from traversing the untreated net. Conversely, and again in line with our prior analysis, the permethrin-treated bed net, with large holes, permitted all life stages to pass through, suggesting that the insecticide did not repel or deter unfed, host-seeking bed bugs.
Table 2 Two-way ANOVA results for blood meal-seeking assays
Blood meal-seeking assays with second instars of two bed bug strains
Because 100% of second instars of the Harlan strain passed through the untreated control bed net (Fig. 5), we compared their responses to those of pyrethroid-resistant bed bugs (Fuller Mill) on all three bed nets. Overall, in all replicates with both strains, ≥ 65% of the nymphs traversed the three bed nets and blood fed (Fig. 6). The two-way ANOVA analysis showed a significant effect of the model on successful blood feeding. All factors assessed, bed bug strain, bed net and the interaction of strain and net significantly impacted blood feeding (Table 2). Throughout the assay Harlan strain bed bugs appeared to be more active than Fuller Mill bed bugs (CCH, personal observation), and a higher percentage of Harlan second instars (96.5%) than Fuller Mill nymphs (85.5%) passed through the untreated bed net and fed (Tukey’s HSD, P = 0.0006) (Fig. 6). The responses of both strains were similar on the permethrin-treated Olyset bed nets, and in line with our prior analysis of net hole length and unfed bed bug width, with 94.0% of Harlan bugs and 90.0% of Fuller Mill bugs feeding (Tukey’s HSD, P > 0.05). Compared to the untreated bed net, lower percentages of second instars of both strains passed through the deltamethrin-treated PermaNet bed nets and fed (Harlan: 72.0%, Tukey’s HSD, P < 0.0001; Fuller Mill: 79.0%, P = 0.0009) (Fig. 6). These findings contrast with the predictions based on our comparison of net hole length and bed bug width, suggesting that deltamethrin may impede passage through the PermaNet bed net. In support of this, we found no significant differences in passage through the untreated net and the permethrin-treated Olyset bed net (Tukey’s HSD, P = 0.2428).
Fig. 6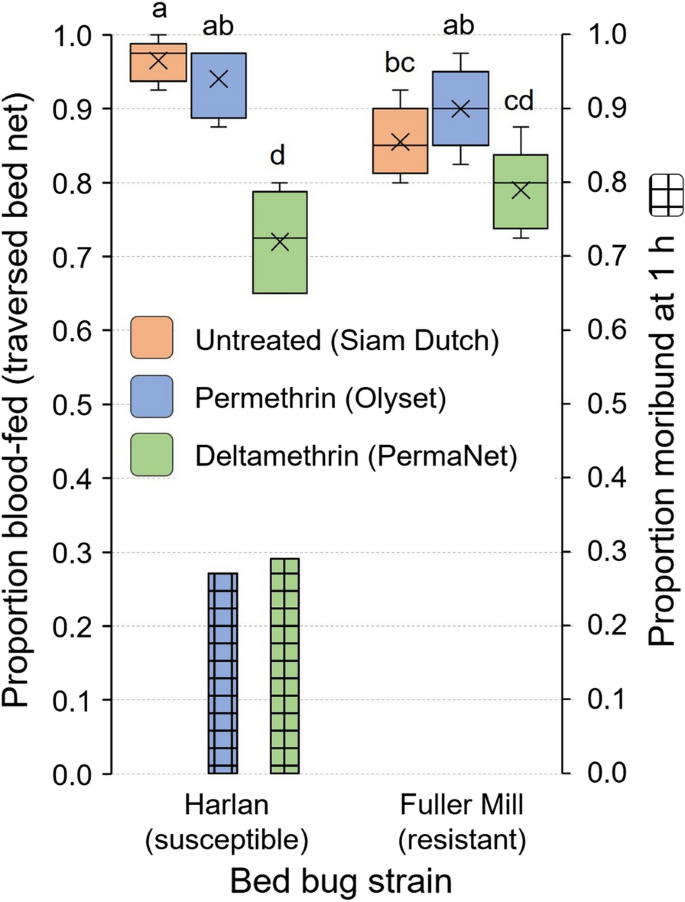
Comparison of the blood meal-seeking assays and mortality of second-instar C. lectularius bed bugs belonging to the insecticide-susceptible Harlan strain and the highly pyrethroid-resistant (resistance ratio > 1000) Fuller Mill strain. Comparisons were made between the two strains, as well as within strains, to assess the impact of pyrethroid resistance on blood meal-seeking behavior. The mean is represented by x within each box plot. Treatments that share lower case letters are not significantly different from each other (ANOVA, Tukey’s HSD, P > 0.05). Significantly lower proportions of blood-fed bed bugs were observed when bed bugs had to traverse deltamethrin-treated (PermaNet) bed nets. At 1 h, some Harlan (susceptible) bed bugs were moribund in these assays, and all moribund bed bugs died at 48 h, whereas no morbidity or mortality was observed in Fuller Mill (resistant) bed bugs
There was no mortality of Harlan or Fuller Mill bed bugs in the 60-min assay with untreated nets. However, 27% and 29% of the Harlan second instars were moribund after the 60-min assays on permethrin- and deltamethrin-treated bed nets, respectively. In contrast, none of the pyrethroid-resistant Fuller Mill bed bugs experienced any morbidity.
Aggregation-seeking assays
The aggregation-seeking assay was designed to simulate the passage of freshly fed bed bugs through a bed net in search of a refuge or aggregation site. We hypothesized that fed and unfed bed bugs would differ in their maneuverability through the three bed nets. This 7-day assay assessed bed bugs as they transitioned from a fully fed state to an unfed state. Across all three bed nets, and throughout the duration of the assay, all fully fed Harlan bed bugs crossed the untreated and permethrin-treated bed nets, and 100% were found in the aggregation jar within 24 h; < 80% crossed the deltamethrin-treated net during the 7-day assay (Fig. 7a). Some Fuller Mill bed bugs failed to cross all three bed nets even after 7 days; thus, < 100% were found in the aggregation jar by the end of the assay (Fig. 7b).
Fig. 7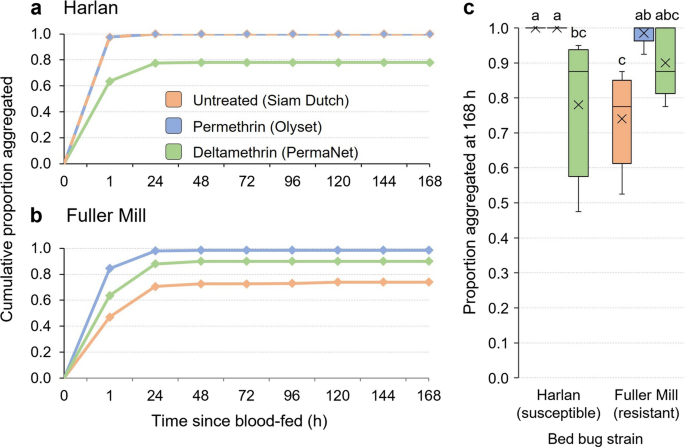
Comparison of cumulative time course of aggregation by second-instar C. lectularius bed bugs belonging to the insecticide-susceptible Harlan strain (a), and highly pyrethroid-resistant (resistance ratio > 1000) Fuller Mill strain (b). Assays were conducted with untreated (Siam Dutch, orange), permethrin-treated (Olyset, blue) and deltamethrin-treated (PermaNet, green) bed nets over the course of 7 days post-feeding or until all the bed bugs traversed the bed net into the aggregation jar. In a, the untreated and permethrin-treated nets followed the same aggregation pattern and so are represented by a dashed line of their respective colors. The majority of all observed aggregation occurred by 48 h for both strains, and Harlan bed bug aggregation was only impeded over the course of this assay by the deltamethrin-treated LLIN. Fuller Mill bed bugs never achieved 100% aggregation through any net, with the untreated net having the largest impeding effect. Statistical comparisons of the cumulative aggregation at 196 h (c). The mean is represented by x within each box plot. Treatments that share lower case letters are not significantly different from each other (ANOVA, Tukey’s HSD, P > 0.05). Significantly fewer Harlan bed bugs passed through the deltamethrin-treated LLIN and aggregated, and significantly fewer Fuller Mill bed bugs passed through the untreated net than the permethrin-treated LLIN
From survival analysis, we estimated the mean aggregation times (time to 50% aggregation) for the Harlan bed bugs as 1.6 ± 0.25, 1.6 ± 0.25 and 14.8 ± 1.38 h through the untreated, permethrin-treated and deltamethrin-treated bed nets, respectively (Fig. 7a). Correspondingly, mean aggregation times for the Fuller Mill nymphs were 33.6 ± 2.84, 5.0 ± 0.71 and 12.3 ± 1.16 h through the same bed nets (Fig. 7b). Cumulative aggregation for both strains across all three nets was compared at 1 h, 24 h and 168 h via two-way ANOVA. The model revealed a significant effect of the interaction between bed net and strain only at 1 h, and significant effects for both the net and strain predictor variables, as well as the interaction between bed net and strain, were seen at 24 and 168 h (Table 3). Overall, there was no significant effect of the permethrin-treated bed net on movement of the Harlan or Fuller Mill nymphs at any of the three time points (Tukey’s HSD tests, P = 0.18, P = 1.00, and P = 1.00, respectively), but a significant effect of the deltamethrin-treated bed net throughout (Tukey’s HSD, P = 0.0001 for all three time points).
Table 3 Two-way ANOVA results for aggregation-seeking assays at 1 h, 24 h and 168 h
Across all three nets the final cumulative aggregation of fed second-instar bed bugs ranged from 47.5% (PermaNet) to 100% (Siam Dutch, Olyset) in Harlan and from 52.5% (Siam Dutch) to 98.5% (Olyset) in Fuller Mill (Fig. 7c). The two-way ANOVA of aggregation at 168 h revealed a significant effect of the overall model on bed bug aggregation (F = 10.64, df = 5, 24, P < 0.001). In fact, bed bug strain, LLIN used and the interaction of strain and LLIN all significantly impacted 168 h aggregation (Table 3). As expected from our prior comparison of net hole length and bed bug width, we found no significant effect of the untreated or permethrin-treated bed net on the 168 h aggregation of Harlan bed bugs (Tukey’s HSD, P = 1.00) (Fig. 7c). However, significantly fewer Harlan nymphs had traversed the deltamethrin-treated net by 168 h (Tukey’s HSD, P = 0.0001), once again suggesting the potential role of a barrier beyond hole size, likely the presence of deltamethrin. In line with our prior statistical comparisons yet again, we found no significant effect of the permethrin- or deltamethrin-treated bed nets on aggregation of Fuller Mill bed bugs at 168 h (Tukey’s HSD, P = 1.00), but unexpectedly, significantly fewer Fuller Mill nymphs traversed the untreated bed net throughout the assay compared to the permethrin-treated bed net (Tukey’s HSD, P = 0.0032) (Fig. 7).
Movement through the untreated bed net toward the aggregation jar caused no mortality in bed bugs of either strain during the 7-day assay (Fig. 8). However, interactions of fully fed Harlan strain nymphs with the permethrin- and deltamethrin-treated bed nets resulted in 2.0 ± 0.9% and 63.5 ± 10.7% mortality, respectively. There was no mortality in the resistant Fuller Mill bed bugs on these two bed nets. Thus, there was no significant difference in mortality of the Harlan and Fuller Mill bed bugs when interacting with the permethrin-treated bed net (Wilcoxon signed-rank test, Z = 1.81, df = 1, P = 0.1667). However, the Harlan nymphs experienced significantly higher mortality than the Fuller Mill bed bugs on deltamethrin-treated bed nets (Wilcoxon signed-rank test, Z = 2.67, df = 1, P = 0.0079) (Fig. 8).
Fig. 8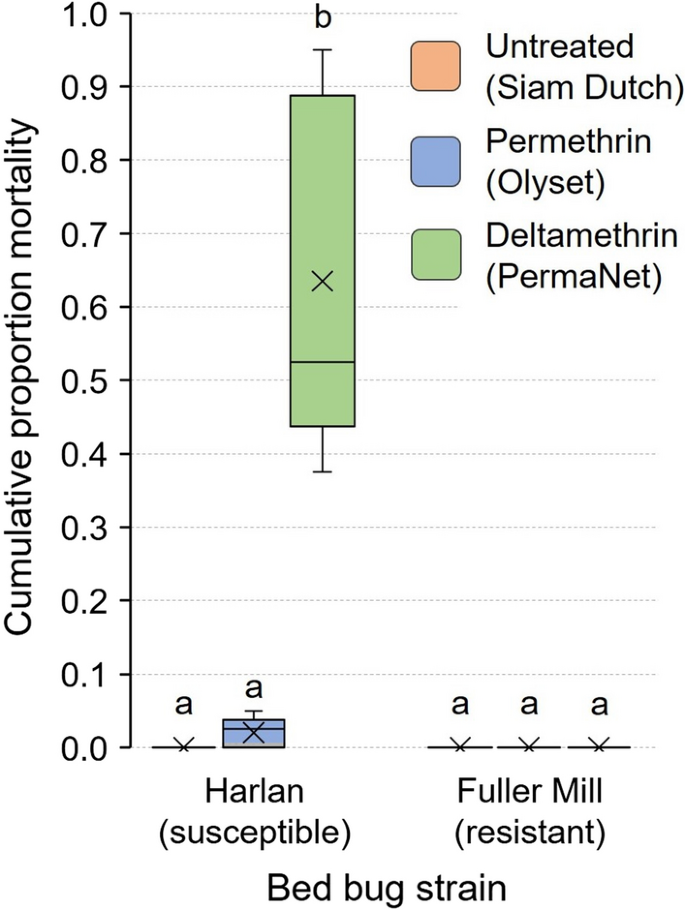
Cumulative 7-day mortality of second-instar C. lectularius bed bugs in the aggregation-seeking assay. Fully fed insecticide-susceptible Harlan strain bed bugs and the highly pyrethroid-resistant (resistance ratio > 1000) Fuller Mill bed bugs were challenged to pass through three different bed nets to reach an aggregation site. The mean is represented by an x within each box plot. Significantly higher mortality in Harlan bed bugs was seen only when they interacted with the deltamethrin-treated LLIN (Wilcoxon signed-rank test, **P = 0.0079)






